Che cos’è l’evoluzione?
Le prime dieci pagine di «Perché l'evoluzione è vera» del biologo Jerry A. Coyne, pubblicato da Codice Edizioni
di Jerry Allen Coyne
Una delle poche certezze che abbiamo sulla natura è il fatto che piante e animali sembrano inestricabilmente e quasi perfettamente progettati per la vita che conducono. I calamari e le sogliole cambiano il colore e il disegno della pelle per confondersi con l’ambiente, diventando invisibili agli occhi di predatori e prede. I pipistrelli hanno una sorta di radar per localizzare gli insetti di notte. I colibrì, che possono librarsi stando fermi in aria e cambiare posizione in un istante, sono molto più agili di un qualsiasi elicottero progettato dall’uomo e hanno una lunga lingua utilissima per succhiare il nettare dai fiori con corolla tubolare. E anche i fiori visitati dai colibrì sembrano appositamente progettati per sfruttare gli uccelli come trait d’union. Infatti, mentre il colibrì è impegnato a bere il nettare, il fiore attacca qualche granello di polline sul suo becco e, in questo modo, riesce a fecondare il fiore successivamente visitato dal colibrì. La natura sembra un ingranaggio ben oliato in cui ogni specie rappresenta una ruota o una qualche molla.
Che cosa potremmo dedurre da tutte queste osservazioni? Di certo l’esistenza di un meccanico esperto. La più famosa esplicitazione di questo ragionamento è quella proposta da William Paley, filosofo inglese del XVIII secolo. Se camminando vediamo un orologio per terra, sosteneva Paley, dovremmo certamente concludere che si tratta dell’opera di un orologiaio. Allo stesso modo, l’esistenza di organismi ben adattati e con caratteri complessi implica l’esistenza di un consapevole progettista divino: Dio. Leggiamo allora l’argomentazione di Paley, una delle più famose nella storia della filosofia:
Quando ci troviamo a ispezionare l’orologio scopriamo […] che le sue diverse parti sono state progettate e assemblate per uno scopo, sono cioè state realizzate e montate insieme per produrre un movimento, un movimento studiato in modo da indicare l’ora del giorno; possiamo inoltre notare che, se le diverse parti avessero una forma diversa da quella che hanno, se avessero una dimensione diversa o se fossero collocate in una posizione diversa o in un qualsiasi ordine diverso da quello che presentano, in quel meccanismo non ci sarebbe nessun movimento o comunque nessuno che potrebbe soddisfare la necessità che adesso soddisfa […]. Qualsiasi segno di un’invenzione, qualsiasi manifestazione di un progetto che si osservi nell’orologio, esiste anche nell’opera della natura, con la differenza, nel caso della natura, di essere molto più grande e più vasto tanto da superare qualsiasi immaginazione.
La tesi sostenua da Paley in modo così eloquente era frutto del buon senso e piuttosto antica. Quando Paley e i suoi seguaci, i sostenitori della “teologia naturale”, descrivevano le piante e gli animali, credevano di catalogare il frutto della grandezza e dell’ingegno di Dio manifestatosi nelle creature tanto straordinariamente ben progettate. Lo stesso Darwin affrontò il tema del progetto divino (prima di liquidarlo) nel 1859. Come si sono potuti sviluppare e perfezionare tutti i finissimi adattamenti di una parte dell’organismo rispetto ad un’altra e alle condizioni di vita e di un organismo rispetto ad un altro organismo? Osserviamo questi mirabili adattamenti reciproci in tutta chiarezza nel picchio e nel vischio e solo un po’ meno chiaramente nei più umili parassiti che aderiscono al pelo di un quadrupede od alle penne di un uccello, nella struttura del coleottero che si immerge nell’acqua, nel seme piumato portato a volo dal più lieve alito di vento. Insomma, osserviamo mirabili adattamenti ovunque ed in tutte le classi del mondo organico. Darwin aveva una propria risposta per risolvere l’enigma della vita. Da appassionato naturalista che studiava a Cambridge (ironicamente occupando le stesse stanze che erano state di Paley) per diventare sacerdote, conosceva bene il potere seduttivo di ragionamenti come quelli di Paley. Più conosciamo le piante e gli animali più ci meravigliamo del modo splendido con cui la loro struttura si adatta alle loro modalità di vita.
Che cosa può esserci di più naturale del pensare che questo adattamento rifletta un progetto conscio? Ma Darwin andò oltre l’ovvio, suggerendo (e sostenendo ciò che affermava con abbondanti prove) due idee che sfatarono per sempre la tesi del progetto deliberato. Queste idee erano la teoria dell’evoluzione e la selezione naturale. Darwin non fu il primo a pensare all’evoluzione, molti l’avevano fatto prima e anche suo nonno Erasmus Darwin aveva vagheggiato l’idea che la vita potesse evolvere. Darwin però fu il primo a dedurre dalla natura le prove in grado di convincere la gente della veridicità dell’evoluzione, mentre la sua idea di selezione naturale era del tutto nuova. Questa intuizione depone a favore del suo genio dato che il concetto di teologia naturale, accettato dalla maggior parte degli eruditi occidentali prima del 1859, è stato del tutto superato nel giro di pochi anni da un solo libro di cinquecento pagine. L’origine delle specie spostò il mistero della diversità della vita dal campo della mitologia a quello della scienza genuina.
E dunque che cos’è il “darwinismo”? La teoria dell’evoluzione per selezione naturale, questa teoria semplice ed estremamente bella, è stata fraintesa così tante volte, e in qualche caso anche deliberatamente, da meritare una veloce rassegna dei suoi punti essenziali. Ritorneremo poi più volte su questi punti quando considereremo le prove che sostengono ognuno di essi. Nella sua sostanza, la moderna teoria dell’evoluzione è facile da capire. Può infatti essere sintetizzata in una sola frase (anche se un po’ lunga): la vita sulla Terra ha subito un’evoluzione graduale a partire da una specie primordiale (forse una molecola in grado di autoreplicarsi) vissuta oltre 3,5 miliardi di anni fa; si è quindi diversificata nel tempo dando origine a molte specie nuove; il meccanismo che sta alla base della maggior parte dei cambiamenti evolutivi (ma non di tutti) è la selezione naturale. Analizzando questa descrizione, vi accorgerete che in realtà è costituita da sei concetti distinti: evoluzione, gradualismo, speciazione, discendenza comune, selezione naturale e cause del cambiamento evolutivo diverse dalla selezione naturale. Vediamo qual è il significato di ciascuno di questi concetti.
La prima idea è l’evoluzione stessa. Questo concetto implica semplicemente che una specie subisce cambiamenti genetici nel corso del tempo. Ciò significa che, nel corso di molte generazioni, una specie può evolvere diventando qualcosa di molto diverso. Le differenze, oggi lo sappiamo, sono basate sui cambiamenti nel dna che si verificano in forma di mutazioni. Le specie di animali e piante attualmente viventi non c’erano in passato, ma discendono da quelle che sono vissute prima. L’uomo, per esempio, si è evoluto a partire da organismi simili a scimmie antropomorfe (ma non identici alle moderne scimmie antropomorfe). Tutte le specie dunque evolvono, ma questo processo non si verifica in ogni specie con la stessa velocità. Il limulo o il gingko quasi non sono cambiati in milioni di anni e infatti la teoria dell’evoluzione non afferma che le specie si evolvono in modo costante né prevede la velocità con cui cambiano. La velocità dipende dalla pressione evolutiva a cui ogni specie è sottoposta. Animali come le balene e gli esseri umani si sono evoluti rapidamente mentre altri, come il celacanto, un noto “fossile vivente”, sono quasi identici per
aspetto ai loro antenati vissuti centinaia di milioni di anni fa.

Il secondo concetto chiave della teoria dell’evoluzione è l’idea di gradualismo. Occorrono molte generazioni per produrre un sostanziale cambiamento evolutivo, come il passaggio dagli uccelli ai rettili. L’evoluzione di nuove caratteristiche, come i denti e le mascelle che distinguono i mammiferi dai rettili, non si verifica nel corso di una o di poche generazioni, ma di solito avviene in centinaia o migliaia (perfino milioni) di generazioni. A onor del vero, è possibile che alcuni cambiamenti si verifichino molto rapidamente.
La durata della vita di una generazione di microrganismi può essere molto ridotta, in alcuni casi occorre soltanto una ventina di minuti per passare da una generazione all’altra. Ciò significa che queste specie possono evolvere e modificarsi anche in modo consistente in un breve intervallo di tempo, come dimostra la comparsa, tristemente rapida, di ceppi di batteri o virus patogeni resistenti ai farmaci. Gli esempi di cambiamenti evolutivi osservabili nel corso della vita di un uomo sono numerosi. Tuttavia quando parliamo di cambiamenti davvero grandi ci riferiamo di solito a trasformazioni che richiedono molte migliaia di anni per verificarsi. Il concetto di gradualismo non implica però che ogni specie evolva sempre con lo stesso ritmo. Proprio come specie differenti evolvono a velocità diversa, così una singola specie evolve più in fretta o più lentamente con l’aumentare o il diminuire della pressione evolutiva. Se la selezione naturale è intensa, per esempio nel caso in cui un animale o una pianta colonizzano un nuovo ambiente, il cambiamento evolutivo può essere rapido. Quando la specie risulta ben adattata a un ambiente stabile, l’evoluzione spesso rallenta.
I due principi seguenti sono come le due facce di una stessa medaglia. È notevole osservare che, pur esistendo molte specie viventi, ognuna di esse (voi, io, l’elefante, il cactus in un vaso) condivide con le altre alcuni tratti fondamentali. Tutte le specie utilizzano per produrre energia un uguale percorso biochimico, tutte sono basate su un codice genetico standard a quattro lettere del dna e questo codice viene letto nello stesso modo per sintetizzare le proteine. Grazie a queste importanti caratteristiche ogni specie può essere fatta risalire a un solo antenato comune, un antenato dotato delle caratteristiche comuni trasmesse ai propri discendenti. Se però l’evoluzione fosse soltanto il cambiamento graduale a livello genetico di una specie, oggi dovrebbe esistere un unico discendente altamente evoluto della specie ancestrale, un’unica specie insomma. In realtà invece le specie sono molte di più: il nostro pianeta oggi ne ospita oltre 10 milioni, tutte diverse, a cui dobbiamo aggiungerne come minimo altre 200 000 fossili. La vita è dunque estremamente diversificata.
Come può tutta questa diversità essere derivata da un’unica forma ancestrale? Qui entra in gioco il terzo concetto fondamentale della teoria dell’evoluzione, l’idea di biforcazione o, più correttamente, di speciazione.
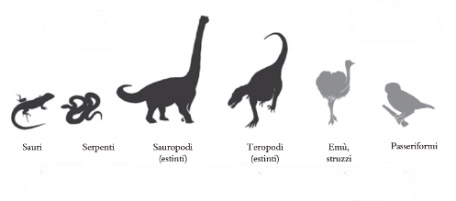
La Figura 1.1 illustra un possibile albero evolutivo dei rettili e degli uccelli. Tutti conosciamo questo tipo di raffigurazioni, ma osserviamo la figura un po’ più in dettaglio per capire che cosa significa effettivamente. Per esempio, che cosa accade esattamente nel punto x corrispondente alla suddivisione che origina da un lato i moderni rettili, come lucertole e serpenti, e dall’altro i moderni uccelli e i loro parenti dinosauri? Il punto (o nodo) x rappresenta una singola specie comune, un antico rettile, che ha prodotto due distinte specie discendenti. Una delle linee di discendenza ha seguito la sua strada biforcandosi varie volte e dando origine a tutti i dinosauri e agli uccelli moderni. L’altra linea di discendenza ha fatto lo stesso producendo la maggior parte dei rettili moderni. L’antenato comune x spesso viene indicato come anello mancante tra i due gruppi discendenti, corrispondente alla connessione genealogica tra gli uccelli e i rettili moderni (l’intersezione che finiremmo per trovare se seguissimo le due linee di discendenza spostandoci indietro nel tempo).

Figura I.I Un esempio di albero evolutivo dei rettili e di discendenza da antenati comuni. x e y sono specie che rappresentano antenati comuni condivisi dalle forme evolutesi più tardi.
Nella Figura 1.1 però è evidenziato anche un secondo anello mancante più recente, il nodo y. Questa specie è l’antenato comune tra dinosauri bipedi e carnivori come il Tyrannosaurus rex (tutti estinti attualmente) e gli uccelli moderni. Anche se gli antenati comuni non sono più qui con noi e i loro fossili sono quasi impossibili da trovare (dopo tutto, rappresentano soltanto una singola specie tra migliaia di altre presenti nella documentazione fossile), qualche volta possiamo scoprire fossili molto vicini a questi antenati, specie provviste di caratteri che dimostrano la discendenza comune. Nel prossimo capitolo, per esempio, scopriremo come erano fatti i dinosauri pennuti che dimostrano l’esistenza del nodo y.
Che cosa accade quando dall’antenato x si ha una biforcazione che dà origine a due specie distinte? Non molto in realtà. Come vedremo più avanti la speciazione implica semplicemente l’evoluzione di gruppi diversi che non possono incrociarsi, vale a dire gruppi tra i quali non possono avvenire scambi di geni. Potendo tornare indietro al tempo in cui la specie che corrisponde all’antenato comune ha incominciato a dividersi, avremmo dunque visto soltanto due popolazioni distinte di questa specie, probabilmente distribuite in luoghi diversi, che incominciavano a evolvere lievi differenze le une rispetto alle altre. Con il passare del tempo (un tempo molto lungo) queste differenze devono essere diventate gradualmente più marcate tanto che i membri delle due popolazioni non hanno più potuto incrociarsi a causa della loro diversità genetica. (Un esito simile può risultare da vari fenomeni: i membri di specie animali diverse possono non riconoscersi più come partner potenziali oppure, se ancora si incrociano tra loro, i discendenti potrebbero essere sterili. Specie differenti di piante, per esempio, possono servirsi di diversi impollinatori oppure fiorire in tempi differenti ostacolando in tal modo la fecondazione incrociata.) Milioni di anni dopo, e dopo altre biforcazioni, una delle specie discendenti di dinosauri, corrispondente al nodo y, si è a sua volta divisa in due specie, una delle quali ha finito per dare origine a tutti i dinosauri carnivori bipedi mentre l’altra ha portato a tutti gli attuali uccelli. Questo momento cruciale della storia evolutiva (la nascita dell’antenato di tutti gli uccelli) non deve essere apparso così straordinario a quel tempo. Non dobbiamo pensare che all’improvviso gli uccelli si siano separati dai rettili, all’epoca del nodo y avremmo semplicemente visto due popolazioni leggermente diverse della stessa specie di dinosauro e questa diversità forse non era neppure più spinta di quella osservabile oggi tra i membri di due differenti popolazioni umane. Tutti i cambiamenti importanti si sono verificati dopo la separazione delle due linee di discendenza, nel corso di migliaia di generazioni, quando la selezione ha agito su una linea promuovendo il volo e sull’altra incentivando i caratteri tipici dei dinosauri bipedi. Soltanto con un’analisi retrospettiva possiamo identificare la specie y come l’antenato comune del T. rex e degli uccelli. Questi lenti episodi evolutivi appaiono di grande importanza soltanto quando ordiniamo in sequenza tutti i discendenti che rappresentano le due linee evolutive divergenti. Ma le specie non devono necessariamente dividersi. La possibilità che ciò accada, come vedremo, dipende dal fatto che le circostanze permettano alle popolazioni di evolvere un numero tale di differenze da non risultare più interfeconde.
La stragrande maggioranza delle specie (oltre il 99% del totale) si estingue senza lasciare alcun discendente. Altre specie, come il gingko, vivono per milioni di anni senza dare origine a nuove specie. La speciazione non è un evento particolarmente frequente, ma ogni volta che una specie si biforca raddoppia il numero delle possibilità che si verifichi una futura speciazione, in tal modo il numero di specie può aumentare esponenzialmente. Nell’arco di periodi molto lunghi della storia, la speciazione, anche se lenta, si verifica abbastanza spesso da poter rendere conto della straordinaria diversità delle piante e degli animali presenti oggi sulla Terra.
Per Darwin era un fenomeno di primaria importanza e infatti decise di inserirla addirittura nel titolo della sua opera più famosa, nella quale sono riportate alcune prove del modo in cui avviene il passaggio da una a più specie. L’unico schema presente in tutta l’Origine è un ipotetico albero evolutivo simile a quello illustrato nella Figura 1.1. Tuttavia Darwin non spiegò davvero come nascono le nuove specie poiché, essendo privo di qualsiasi nozione di genetica, non sapeva che per rendere ragione dell’origine delle specie occorre scoprire come si formano le barriere in grado di impedire gli scambi genetici. Una prima spiegazione di come avviene la speciazione è stata pertanto proposta soltanto negli anni Trenta del Novecento. Dirò di più su questo tema, che tra l’altro è il mio campo di ricerca, nel Capitolo 7. Se la storia della vita può essere esemplificata da un albero, con tutte le specie che si originano a partire da un singolo tronco, è logicamente possibile scoprire un’origine comune per ciascuna coppia di ramificazioni (corrispondenti alle specie esistenti) seguendo ogni ramificazione all’indietro finché non si giunge all’intersezione nel ramo, più grande, comune. Questo nodo, come abbiamo visto, è rappresentato dall’antenato comune delle due specie da cui siamo partiti. Seguendo il percorso inverso, dal passato al presente, se la vita incomincia con una specie e si divide in milioni di specie discendenti attraverso un processo di ramificazione, ne consegue che ciascuna coppia di specie deve avere in comune un antenato nel suo passato. Le specie, così come le persone, strettamente imparentate hanno un antenato comune che è vissuto piuttosto di recente, mentre l’antenato comune delle specie, e delle persone, imparentate più alla lontana è vissuto in un passato più remoto. Per questo motivo l’idea di discendenza comune (il quarto concetto chiave del darwinismo) rappresenta l’altra faccia della medaglia rispetto alla speciazione. Tale idea implica che possiamo sempre guardare indietro nel tempo, servendoci delle sequenze del dna e/o dei fossili, e trovare linee di

Figura I.2 Una filogenesi (albero evolutivo) dei vertebrati, che rivela come l’evoluzione produce un raggruppamento gerarchico dei caratteri, e dunque di specie che presentano quei caratteri. I punti indicano dove compare sull’albero ciascun carattere considerato.
discendenza che si riuniscono convergendo nel loro antenato comune. Esaminiamo l’albero evolutivo dei vertebrati (Figura 1.2). Nella figura ho riportato alcune delle caratteristiche usate dai biologi per dedurre le relazioni evolutive. Per cominciare, pesci, anfibi, mammiferi e rettili hanno tutti una colonna vertebrale (sono tutti “vertebrati”) perciò devono essere derivati tutti da un antenato comune che, a sua volta, aveva le vertebre. Ma all’interno del gruppo dei vertebrati i rettili e i mammiferi condividono un altro carattere chiave: diversamente dai pesci e dagli anfibi, hanno infatti un uovo amniotico, cioè provvisto di una sacca membranosa piena di fluido, detta amnios, che avvolge l’embrione proteggendolo.
Di conseguenza rettili e mammiferi devono condividere un antenato comune più recente rispetto agli altri vertebrati, un antenato che possedeva un uovo amniotico. Questo sottogruppo di vertebrati con uovo amniotico è divisibile ulteriormente in due sottogruppi, uno con specie tutte provviste di pelo, omeoterme e che allattano i piccoli (si tratta dei mammiferi) e l’altro con specie eteroterme, provviste di squame e che depongono uova con guscio (cioè i rettili). Come tutte le specie anche quelle dei vertebrati formano una gerarchia ramificata, in cui grandi gruppi costituiti da specie che condividono pochi tratti sono suddivisi in gruppi più piccoli, di specie che hanno in comune ancora più caratteristiche, e così via fino ad arrivare alle singole specie, come il baribal e il grizzly che condividono quasi tutte le loro caratteristiche.
A dire il vero l’organizzazione ramificata della vita è stata riconosciuta molto prima di Darwin. Fin dal 1735, quando il botanico svedese Carl von Linné (Carolus Linnaeus) pubblicò il Systema Naturae, i biologi incominciarono a classificare gli animali e le piante in modo sistematico scoprendo che esse si inserivano bene in una classificazione che veniva definita naturale. Sorprendentemente, infatti, biologi diversi finivano per suggerire raggruppamenti quasi identici, che, di conseguenza, non erano artefatti soggettivi del bisogno umano di classificare, ma rappresentavano la diretta conseguenza di un aspetto della natura fondamentale quanto vero. Tuttavia nessuno ha saputo dire di che cosa si trattasse finché non è arrivato Darwin, il quale ci ha spiegato che l’organizzazione ramificata della vita è precisamente quanto l’evoluzione prevede. Gli organismi con antenati comuni recenti condividono molti tratti, mentre quelli con antenati comuni vissuti in un passato più lontano sono meno simili. La classificazione naturale è, di per sé, una valida prova dell’evoluzione. Perché? Perché se cerchiamo di mettere in ordine oggetti che non sono derivati da un processo evolutivo di divisione e discendenza non vediamo una simile organizzazione ramificata.
Consideriamo, per esempio, le scatole di fiammiferi della mia collezione. Queste scatole non si prestano a una classificazione naturale come le specie viventi. Possiamo ordinarle gerarchicamente basandoci sulla dimensione e poi suddividerle in base al paese d’origine, quindi in base al colore, nell’ambito del singolo paese, e così via. In alternativa si potrebbe considerare il disegno sulla scatola, passando poi per il colore e quindi per la data. Esistono molti modi per riordinare le scatole di fiammiferi e ognuno può farlo in modo diverso. Ovviamente non esiste un sistema sul quale tutti i collezionisti possono concordare perché ciascuna scatola di fiammiferi non si è evoluta dando origine a un’altra scatola lievemente differente, ma è stata progettata da zero in base al capriccio umano. Le scatole di fiammiferi sarebbero simili agli esseri viventi se gli organismi fossero il frutto di una creazione. In questo caso i viventi non avrebbero una discendenza comune, ma sarebbero semplicemente il frutto di una creazione istantanea di forme pensate ex novo per adattarsi al proprio ambiente. Se le cose stessero così non potremmo aspettarci di vedere una gerarchia ramificata delle specie riconosciuta da tutti i biologi. Fino a trent’anni fa circa i biologi si basavano su caratteri visibili come l’anatomia e le modalità di riproduzione per ricostruire la discendenza delle specie viventi. Questo metodo era basato sul presupposto per cui organismi con caratteri simili hanno anche geni simili e, di conseguenza, sono più strettamente correlati.
Ora però siamo in possesso di un mezzo nuovo, potente e indipendente, per stabilire la discendenza: possiamo infatti esaminare e confrontare direttamente i geni. Sequenziando il dna di varie specie e misurando il grado di somiglianza tra queste sequenze possiamo ricostruire le relazioni evolutive tra le specie. Per applicare questo metodo si parte dal ragionevole presupposto per cui le specie con dna più simili hanno una parentela più stretta, cioè più due specie si somigliano più il loro antenato comune deve essere recente. Le analisi molecolari non hanno prodotto radicali cambiamenti negli alberi evolutivi realizzati prima degli studi sul dna: di solito i tratti visibili degli organismi e le sequenze dei loro dna forniscono le stesse informazioni riguardo le parentele in termini evolutivi. L’idea della discendenza comune ci permette di avanzare alcune previsioni scientifiche importanti e verificabili sull’evoluzione. Se gli uccelli e i rettili possono essere raggruppati insieme sulla base dei loro caratteri anatomici e delle sequenze di dna, possiamo prevedere di trovare nella documentazione fossile antenati comuni di entrambi. Questa previsione scientifica è stata ampiamente soddisfatta, fornendo così una delle prove più forti a sostegno dell’evoluzione. Incontreremo alcuni di questi antenati nel prossimo capitolo.
Il quinto concetto fondamentale della teoria dell’evoluzione fu chiaramente considerato da Darwin come il suo più grande contributo intellettuale: si tratta infatti della selezione naturale. In realtà non fu l’unico a proporre questa idea (come è noto, il suo contemporaneo, il naturalista Alfred Russel Wallace, propose più o meno la stessa idea dando luogo a uno dei più famosi esempi di scoperte contemporanee ma indipendenti della storia della scienza), tuttavia Darwin si guadagnò la fetta maggiore del merito perché nell’Origine sviluppò il concetto di selezione in grande dettaglio, fornendo numerose prove a favore e analizzandone le conseguenze principali. La selezione naturale, che al tempo di Darwin era la parte della teoria dell’evoluzione considerata più rivoluzionaria, ancora oggi sembra disturbare molti. L’idea è rivoluzionaria proprio per lo stesso motivo che la rende disturbante: spiega infatti il progetto apparentemente insito nella natura come un processo puramente materialistico che non richiede un evento creativo né la guida da parte di forze soprannaturali.
La selezione naturale non è difficile da capire. Se gli individui che appartengono a una specie differiscono uno dall’altro dal punto di vista genetico e se alcune di queste differenze influenzano la capacità di un individuo di sopravvivere e riprodursi nel proprio ambiente, allora, nella successiva generazione, i geni “buoni” che portano a una maggiore probabilità di sopravvivere e riprodursi saranno in numero superiore rispetto ai geni “non altrettanto buoni”. Con il passare del tempo la popolazione diventa gradualmente sempre più adatta al proprio ambiente dato che le mutazioni utili emergono e si diffondono nella popolazione, mentre al contrario quelle dannose vengono estirpate.
Questo processo finisce quindi per produrre organismi che sono ben adattati ai vari ambienti e alle diverse modalità di vita. Vediamo un esempio. Il mammut viveva nelle regioni settentrionali dell’Eurasia e dell’America settentrionale ed era adattato al freddo perché aveva una folta pelliccia di lunghi peli (esemplari completi e congelati sono stati trovati sepolti nella tundra). Questa specie discendeva probabilmente da antenati con poco pelo e aspetto simile a quello dei moderni elefanti. Le mutazioni verificatesi nella specie ancestrale hanno portato alla comparsa, tra i mammut, di singoli individui più pelosi (come peraltro si verifica nella nostra specie).
In seguito al raffreddamento del clima, o alla diffusione della specie nelle aree più settentrionali, gli individui più pelosi si sono rivelati avvantaggiati nell’ambiente freddo e sono riusciti a lasciare più discendenti rispetto ai mammut meno pelosi. Così la popolazione si è arricchita di geni per il pelo folto. Nella generazione successiva, i mammut in media saranno stati più pelosi rispetto a quelli della generazione precedente. Lasciate che questo processo continui per qualche migliaio di generazioni e vedrete i mammut glabri di partenza rimpiazzati da quelli pelosi. Se caratteri diversi dovessero poi rivelarsi altrettanto utili per sopravvivere al freddo (per esempio, la dimensione del corpo, la quantità di grasso e così via) vedremo che anch’essi si modificheranno, proprio come la pelliccia.
Il processo è davvero semplice e richiede soltanto che gli individui di una specie presentino una variabilità genetica in grado di influenzare la loro capacità di sopravvivere e di riprodursi nell’ambiente in cui vivono. Se questo accade, la selezione naturale (e l’evoluzione) è inevitabile. Come vedremo un tale requisito è soddisfatto in tutte le specie esaminate. Considerato inoltre che molti tratti possono influenzare l’adattamento di un individuo al proprio ambiente (la cosiddetta fitness), è chiaro che la selezione naturale può davvero modellare un animale o una pianta, con il passare degli eoni, dando origine a qualcosa che sembra il frutto di un progetto deliberato. È importante capire che esiste una profonda differenza tra ciò che ci dovremmo aspettare di vedere se gli organismi fossero frutto di un progetto conscio e quello che potremmo vedere se fosse (come è) la selezione naturale a influenzare l’evoluzione delle specie.
La selezione naturale non è un ingegnere esperto ma ricorda di più una sorta di “trafficone”. Non eguaglia la perfezione raggiungibile da un ingegnere a partire da un progetto, ma produce il meglio che può con gli elementi a disposizione. Le mutazioni utili per produrre un progetto perfetto potrebbero non emergere perché sono semplicemente troppo rare. I rinoceronti africani, con i loro due corni, possono risultare meglio adattati a difendersi e a sfidare i propri compagni rispetto al rinoceronte indiano a cui è stato concesso soltanto un corno (in realtà non si tratta di veri corni, ma di peli fusi insieme). Tuttavia è possibile che una mutazione in grado di produrre due corni semplicemente non si sia verificata nei rinoceronti indiani. D’altra parte un corno è comunque meglio che non averne affatto e il rinoceronte indiano è comunque meglio del suo antenato senza corni, insomma nella storia genetica della specie il caso potrebbe aver promosso un “progetto” non esattamente perfetto. E naturalmente qualsiasi esempio di pianta o di animale attaccato da un parassita o colpito da malattia rappresenta un insuccesso nell’adattamento. Lo stesso discorso vale per tutti i casi di estinzione, che rappresentano ben oltre il 99% delle specie comparse sulla Terra. Questo particolare, tra l’altro, costituisce un enorme problema per la teoria del disegno intelligente. Non sembra infatti così intelligente progettare milioni di specie destinate a estinguersi per poi sostituirle con altre simili che, nella maggior parte dei casi, a loro volta scompariranno. I sostenitori del disegno intelligente però non hanno mai affrontato davvero questa difficoltà.
***
È uscito per Codice Edizioni Perché l’evoluzione è vera, di Jerry Allen Coyne.
Coyne è un biologo americano, noto per il suo contributo al dibattito sulla teoria del “disegno intelligente”, e per le sue posizioni secondo cui religione e scienza sono profondamente incompatibili. Attualmente è docente presso l’Università di Chicago, presso il Dipartimento di Ecologia ed Evoluzione.